
Этот цикл статей описывает волновую модель мозга, серьезно отличающуюся от традиционных моделей. Настоятельно рекомендую тем, кто только присоединился, начинать чтение с первой части.
Мы исходим из того, что явления внешнего мира воздействуют на наши органы чувств, вызывая определенные потоки сигналов в нервных клетках. В процессе обучения кора приобретает способность детектировать определенные сочетания сигналов. Детекторами выступают нейроны, синаптические веса которых настраиваются на картины активности, соответствующие детектируемым явлениям. Нейроны коры следят за своим локальным окружением, образующим их локальное рецептивное поле. Информация на рецептивные поля нейронов поступает либо посредством топографической проекции, либо через распространение волн идентификаторов, несущих уникальные узоры, соответствующие уже выделенным признакам. Нейроны-детекторы, реагирующие на одно и то же сочетание признаков, образуют детекторные паттерны. Узоры этих паттернов определяют уникальные волны идентификаторов, которые эти паттерны запускают, приходя в состояние вызванной активности.
В 1952 году Алан Тьюринг опубликовал работу под названием «Химические основы морфогенеза» (Turing A. M., 1952), посвященную самоорганизации материи. Сформулированный им основной принцип гласил, что глобальный порядок определяется локальным взаимодействием. То есть, чтобы получить структурную организацию всей системы, необязательно иметь некий глобальный план, а можно ограничиться исключительно заданием правил близкого взаимодействия образующих систему элементов.
Обучение нейронов происходит не изолированно, а с учетом активности их окружения. Правила учета этой активности определяют самоорганизацию коры. Самоорганизация означает, что по ходу обучения не просто формируются детектирующие нейронные паттерны, а что эти паттерны выстраиваются в некие пространственные конструкции, имеющие свой определенный смысл.
Наиболее очевидный способ самоорганизации – это структурирование по степени близости. Можно обучать нейроны так, чтобы паттерны, соответствующие близким в некотором смысле понятиям, оказывались рядом и в пространстве коры. Позже мы увидим, что такое размещение окажется необходимым для реализации многих важных функций, свойственных мозгу. Пока же просто посмотрим на механизмы, которые могут обеспечить такую организацию.
Достаточно неоднозначный вопрос – как мерить близость понятий. Один подход связан с тем, что любое понятие можно сопоставить с неким описанием. Например, описанием события может быть вектор, компоненты которого показывают выраженность в событии определенных признаков. Набор признаков, в которых ведется описание, образует описательный базис. В таком случае, разумная мера близости – это близость описаний. Чем ближе описание двух явлений, тем более похожи друг на друга эти явления. В зависимости от задачи, которая далее решается с использованием меры близости, можно выбрать тот или иной алгоритм расчета. Мера близости тесно связана с понятием расстояния между объектами. Одно можно пересчитать в другое и наоборот.
Иной подход – это временная близость явлений. Если два явления происходят одновременно, но при этом имеют разные описания, то все равно можно говорить об их определенной близости. Чем чаще явления случаются совместно, тем сильнее может быть истолкована их близость.
На самом деле эти два подхода не противоречат, а взаимодополняют друг друга. Если два явления часто случаются совместно, то надо сделать вывод о наличии явления, которое является реализацией этой совместности. Это новое явление само может быть признаком, описывающим оба исходных явления. Теперь у этих явлений появилось общее в описании, а значит, и близость в описательном подходе. Будем называть близость, учитывающую одновременно и описательную и временную близость, обобщенной близостью.
Представьте, что вы разглядываете какой-либо предмет с разных сторон. Возможно, эти виды будут совершенно различны в описательном плане, но тем не менее нельзя отрицать их близость в некотором ином смысле. Временная близость – это возможное проявление этого иного смысла. Несколько позже мы значительно глубже разовьем эту тему, сейчас же просто отметим необходимость учитывать оба подхода при определении близости понятий.
Наиболее простая для понимания нейронная сеть, которая хорошо иллюстрирует самоорганизацию по степени описательной близости – это самоорганизующиеся карты Кохонена (рисунок ниже).

Пусть у нас есть входная информация, заданная вектором
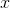 . Есть двумерная решетка из нейронов. Каждый нейрон связан с входным вектором , эта связь определяется набором весов
. Есть двумерная решетка из нейронов. Каждый нейрон связан с входным вектором , эта связь определяется набором весов  . Вначале инициируем сеть случайными малыми весами. Подавая входной сигнал, для каждого нейрона можно определить его уровень активности как линейного сумматора. Возьмем нейрон, который покажет наибольшую активность, и назовем его нейроном-победителем. Далее сдвинем его веса в сторону образа, на который он оказался похож. Более того, проделаем аналогичную процедуру для всех его соседей. Будем ослаблять этот сдвиг по мере удаления от нейрона-победителя.
. Вначале инициируем сеть случайными малыми весами. Подавая входной сигнал, для каждого нейрона можно определить его уровень активности как линейного сумматора. Возьмем нейрон, который покажет наибольшую активность, и назовем его нейроном-победителем. Далее сдвинем его веса в сторону образа, на который он оказался похож. Более того, проделаем аналогичную процедуру для всех его соседей. Будем ослаблять этот сдвиг по мере удаления от нейрона-победителя.=\omega _{j}(n)+\eta(n) h_{j,i(x)}(n)(x-\omega _{j}(n)))
здесь –
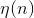 скорость обучения, которая падает со временем,
скорость обучения, которая падает со временем,  — амплитуда топологической окрестности (зависимость от n предполагает, что она тоже со временем уменьшается).
— амплитуда топологической окрестности (зависимость от n предполагает, что она тоже со временем уменьшается).Амплитуда окрестности может быть выбрана, например, Гауссовой функцией:
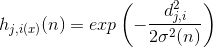
где
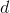 – расстояние между корректируемым нейроном
– расстояние между корректируемым нейроном 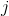 и нейроном-победителем
и нейроном-победителем 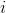 .
.
Функция Гаусса
По мере обучения на такой самоорганизующейся карте будут выделяться зоны, соответствующие тому, как распределены обучающие образы. То есть сеть сама определит, когда во входном потоке встретятся похожие друг на друга картины, и создаст для них близкие представительства на карте. При этом, чем сильнее будут различаться образы, тем более обособленно друг от друга будут расположены и их представительства. В итоге, если соответствующим образом раскрасить результат обучения, то он будет выглядеть примерно так, как показано на рисунке ниже.

Результат обучения карты Коханена
Карты Коханена используют функцию амплитуды топологической окрестности, что предполагает, что нейроны кроме взаимодействия через синапсы могут обмениваться дополнительной информацией, говорящей о характере окружающей активности, и эта информация может влиять на ход их синаптического обучения. Необходимость в передаче дополнительной информации возникала несколько раньше и в нашей модели. Описывая обучение внесинаптических рецепторов, мы вводили правила, опирающиеся на знание нейронами окружающей активности определенного типа. Например, знание общего уровня активности позволяло нам принимать решения как о необходимости обучения, так и об отказе от него.
Чтобы оставаться при построении модели в рамках определенной биологической достоверности, попробуем показать, какие механизмы в реальной коре могут отвечать за расчет и передачу дополнительной незакодированной в сигналы аксонов информации.
Около 40 процентов объема мозга занимают глиальные клетки. Их общее количество где-то на порядок превосходит количество нейронов. Традиционно на глиальные клетки возлагается масса сервисных функций. Они создают объемный каркас, заполняя пространство между нейронами. Участвуют в поддержании гомеостаза среды. В процессе развития нервной системы задействованы для формирования топологии мозга. Шванновские клетки и олигодендроциты отвечают за миелинизацию крупных аксонов, что приводит к многократному ускорению передачи нервных импульсов.
Так как глиальные клетки не генерируют потенциалов действия, они не участвуют напрямую в информационном взаимодействии. Но это не значит, что они полностью лишены информационных функций. Например, плазматические астроциты располагаются в сером веществе и имеют многочисленные сильно ветвящиеся отростки. Эти отростки опоясывают окружающие синапсы и влияют на их работу (рисунок ниже).

Астроцит и синапс (Филдз, 2004)
Так, например, был описан следующий механизм (R. D. Fields, B. Stevens-Graham, 2002). Активация нейрона ведет к тому, что из его аксона высвобождаются молекулы АТФ. АТФ (аденозинтрифосфат) — нуклеотид, играющий исключительно важную роль во всем организме, основная его функция – обеспечение энергетических процессов. Но кроме этого АТФ способен выступать и как сигнальное вещество. Под его воздействием инициируется перемещение кальция внутрь астроцита. Это, в свою очередь, приводит к тому, что астроцит высвобождает собственный АТФ. В результате происходит передача такого состояния на соседние астроциты, которые передают его еще дальше. При этом поглощение кальция астроцитом ведет к тому, что он начинает влиять на синапсы, с которыми контактирует. Астроциты способны как усиливать реакцию синапса за счет выброса соответствующего медиатора, так и ослаблять ее за счет его поглощения или выброса связывающих нейромедиатор белков. Кроме того, астроциты способны выделять сигнальные молекулы, регулирующие выброс медиатора аксоном. Концепция передачи сигналов между нейронами, учитывающая влияние астроцитов, называется трехсторонним синапсом.
Такое взаимодействие астроцитов и нейронов не передает конкретных информационных образов, но оно очень хорошо подходит на роль механизма, обеспечивающего распространение «поля активности», которое может управлять обучением синапсов и таким образом задавать пространственные координаты для новых детекторных паттернов.
Кроме астроцитов на поведение синапсов влияет еще и межклеточный матрикс. Матрикс – это множество молекул, произведенных клетками мозга и заполняющих межклеточное пространство. В статье (Dityatev A., Schachner M., Sonderegger P., 2010) показано, что изменение состава матрикса влияет на характер синаптической пластичности, то есть на обучение нейронов.
Импульсная активность нейронов создает точечный пространственный узор, который динамично меняется, кодируя информационные потоки. Эта активность изменяет состояние окружающей глиальной среды и среды матрикса таким образом, что создается нечто наподобие поля обобщенной активности (рисунок ниже).

Точечная активность и поле активности
Поле активности, с одной стороны, размывает точечную активность, создавая область, простирающуюся за пределы активных нейронов, с другой стороны, обладает инерционностью и продолжает существовать какое-то время после прекращения импульсной активности.
Если внутри этого поля активности мы создадим детектор текущего подаваемого образа, то он окажется поблизости от похожих на него детекторов. При этом похожесть может оказаться как похожестью по рецептивному полю, так и похожестью, возникающей за счет совмещения событий по времени.
В реальном мозге лучше всего пространственная организация изучена для зоны первичной зрительной коры. За счет преобразований, начинающихся еще в сетчатке, на первичную кору поступает сигнал, в котором основная информация – это линии, описывающие контуры объектов на исходном изображении. Нейроны первичной зрительной коры видят, в основном, небольшие фрагменты этих линий, проходящие через их рецептивные поля. Неудивительно, что существенная часть нейронов этой зоны – это детекторы линий, идущих под различными углами.
Экспериментально выявлено, что нейроны, располагающиеся вертикально друг под другом, реагируют на один и тот же стимул. Группу таких нейронов принято называть кортикальной мини колонкой. Вернон Маунткасл (В. Маунткасл, Дж. Эдельман, 1981) выдвинул гипотезу, что для мозга кортикальная колонка – это основная структурная единица переработки информации.
Мы ранее, говоря о паттернах нейронов детекторов, изображали их группами нейронов, распределенных в некой локальной области. Это было следствием того, что при моделировании, и соответственно, подготовке картинок использовались плоские нейронные сети. Реальная кора трехмерна. Объемность коры не влияет на наши рассуждения о возникновении и распространении волн идентификаторов. В трехмерной коре волны распространяются точно также как и в плоской. Но в объемной коре ничто не мешает нам, расположить нейроны-детекторы, образующие единый паттерн, вертикально друг под другом. Такое расположение ни хуже и не лучше любых других. Главное требование к паттерну – это случайность его узора. Так как связи нейронов распределены случайным образом, то расположенные вертикально в одной кортикальной колонке нейроны вполне можно считать случайным паттерном. Такое вертикальное расположение достаточно удобно при определении места создания детекторного паттерна. Не надо определять локальную область, а достаточно указать позицию, в которой требуется создать паттерн. Выбор позиции может быть, например, там, где поле активности максимально для всех свободных колонок некой окрестности. Можно предположить, что кортикальные миниколонки реальной коры – это и есть детекторные паттерны, описываемые в нашей модели.
Для традиционных моделей коры, не учитывающих волновые сигналы, объяснение того, что все нейроны миниколонки реагируют на один и тот же стимул, представляет определенное затруднение. Приходится предполагать, что это либо дублирование для обеспечения отказоустойчивости, либо попытка собрать вместе нейроны, реагирующие на один стимул, но настроенные на его определение в разных позициях общего рецептивного поля. Последнее обыгрывалось в неокогнитроне через использование плоскостей простых клеток. Волновая модель позволяет взглянуть на это с иной стороны. Можно предположить, что задача кортикальной мини колонки – это, узнав характерный стимул, запустить соответствующий волновой идентификатор. Тогда несколько десятков нейронов, образующих мини колонку и реагирующих совместно, — это и есть механизм запуска волны, то есть детекторный паттерн.
Организация ориентационных колонок в реальной зрительной коре образует так называемые «волчки» («pinwheel»). Такой «волчок» (рисунок ниже (B)) имеет центр, куда сходятся колонки разных ориентаций, и расходящиеся хвосты, для которых характерна плавность смены предпочитаемого стимула. Один волчок образует гиперколонку. Хвосты разных гиперколонок переходят друг в друга, образуя ориентационную карту коры (рисунок ниже (A)).

Распределение ориентационных колонок в реальной коре, полученное оптическим методом (Николлс Дж., Мартин Р., Валлас Б., Фукс П., 2003)
На примере зрительной коры удобно сравнить реальное распределение ориентационных колонок и результаты моделирования с использованием поля активности.
Будем подавать на фрагмент коры линии под различными углами. Активность нейронов-детекторов будем считать, как косинус угла между поданным образом и образом на детекторе.

Нейрон с индексом
имеет координаты на коре  . Значение поля активности
. Значение поля активности  от нейрона в точке с координатами
от нейрона в точке с координатами 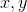 можно представить, например, через Гауссово распределение:
можно представить, например, через Гауссово распределение:
В каждой точке коры итоговое значение поля активности можно записать:
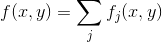
Будем размещать новые детекторы в такой свободной позиции, для которой значение максимально. Результат такого обучения показан на рисунке ниже.

Результат обучения детекторов (слева), поле активности (справа)
Как видно, результат моделирования очень похож на реальное распределение. Но надо отметить, что приведенный пример очень условен. В реальной коре каждая колонка несколько смещена относительно соседей и следит за немного другим фрагментом изображения. Это создает «хвосты» из детекторов, настроенных на одну ориентацию. Но при этом стимулы таких детекторов – это разные образы, основанные на разных рецептивных полях.
Пространственную самоорганизацию коры можно сопоставить с самоорганизацией материи в окружающем нас мире. Атомы, молекулы, предметы, планеты, звезды, галактики, вселенная – все это следствие существования четырех фундаментальных взаимодействий. Принято говорить о гравитационном, электромагнитном, сильном и слабом взаимодействии. Частицы материи создают вокруг себя поля, соответствующие взаимодействиям. Однотипные поля суммируются и формируют результирующее поле. Результирующие поля оказывают влияние на частицы, определяя их поведение. Где-то так же обстоит дело и с мозгом. Похоже, что кора может создавать несколько типов полей, обладающих разными свойствами. Каждое из полей оказывает свое воздействие на поведение нейронов. Совокупность этих взаимодействий, например, может определять алгоритм обучения синапсов и тем самым пространственную организацию нейронов-детекторов.
Нетрудно заметить, что пространственная группировка по обобщенной близости содержит внутреннее противоречие. В связи с этим вспоминается анекдот. Решил как-то лев поделить зверей на красивых и умных. И вот стоит растерянная обезьяна: «И что мне теперь, разорваться?». Сложности возникают, когда размещаемое понятие оказывается близко к другим, удаленным друг от друга понятиям. Например, если мы раскладываем книги по полкам библиотеки, то нам одновременно хочется сгруппировать их и по жанрам, и по авторам, и по языку оригинала, и по читательскому рейтингу. Но если книга только одна, то возникает определенная проблема.
В процессе самоорганизации коры может оказаться, что одно и то же понятие близко в обобщенном смысле сразу к нескольким другим, расположенным в разных местах коры. Не имеет смысла размещать детекторный паттерн компромиссно, пытаясь найти некое равноудаленное место. Во-первых, такого места может не быть, а во-вторых, это полностью нарушит принцип «похожее рядом». Не остается ничего другого, как создать несколько детекторных паттернов, каждый поблизости от своего локального максимума поля активности.
На первый взгляд, дублирование одного и того же понятия в разных местах выглядит некрасиво. Работая с информацией, всегда хочется прийти к ее интеграции, а не распылению. Но наша модель нейронной сети позволяет достаточно изящно снять возникшие противоречия. Каждый из детекторных паттернов, хотя и создается в разных местах коры, обучается на одну и ту же волну идентификаторов. При этом, если брать двухуровневую конструкцию, один из уровней фиксирует на себе фрагмент узора волны общего идентификатора. Это означает, что разбросанные по разным местам детекторные паттерны оказываются связаны между собой в единое понятие с общим идентификатором.
Такое поведение понятий – следствие задекларированного ранее дуализма. Понятие – это одновременно и паттерн и волна. Паттерны порождают волну, волна активирует паттерны. Пространственное положение паттернов, относящихся к одному понятию, характеризует понятие через его близость к другим понятиям. Положение паттернов на коре, по сути, очень многое говорит о свойствах и характеристиках соответствующего явления. Но если в картах Кохонена одному понятию соответствует одна область, как получается из парадигмы «победитель забирает все», то в нашей модели удается не только дать более богатое описание, но за счет дуализма еще и сохранить единство составляющих.
Самоорганизация по принципу «похожее рядом» не является для коры самоцелью. Позже мы покажем, что важнейшие для работы мозга алгоритмы требуют именно такой организации и практически нереализуемы вне ее.
А теперь не совсем банальная физическая аналогия. Основная идея квантовой физики заключается в том, что квантовая система может находиться не в любых возможных состояниях, а только в некоторых, допустимых для нее. При этом квантовая система не принимает никакого определенного значения, пока не произойдет измерение. Только в результате измерения система переходит в одно из разрешенных ей состояний. Под измерением понимается любое внешнее взаимодействие, которое заставляет квантовую систему проявить себя.
В квантовой физике вводится комплекснозначная функция, описывающая чистое состояние объекта, которая называется волновой функцией. В наиболее распространенной копенгагенской интерпретации эта функция связана с вероятностью обнаружения объекта в одном из чистых состояний (квадрат модуля волновой функции представляет собой плотность вероятности). Гамильтоновой системой называется динамическая система, описывающей физические процессы без диссипации. Пока квантовая система не вступит во взаимодействие, она является гамильтоновой. Описать поведение такой системы во времени и пространстве можно, описав эволюцию ее волновой функции. Эта эволюция определяется уравнением Шредингера.
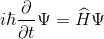
где –
 гамильтониан – оператор полной энергии системы. Его спектр – это множество возможных состояний, в которых система может оказаться после измерения.
гамильтониан – оператор полной энергии системы. Его спектр – это множество возможных состояний, в которых система может оказаться после измерения.Когда происходит измерение, квантовая система принимает одно из разрешенных состояний. Это называется редукцией или коллапсом волновой функции. В процессе измерения система перестает быть изолированной, и ее энергия может не сохраняться, так как происходит обмен энергией с прибором.
Какое состояние примет квантовая система в момент измерения – дело случая. Но вероятность выбора каждого из состояний не одинакова, она определяется связанным с этим состоянием значением волновой функции.
В нашей модели детекторные паттерны создаются в позициях коры, где наблюдается описательная или временная близость нового понятия и понятий уже существующих. Тем самым формируется набор мест, описывающих вероятность проявления понятия в той или иной ситуации. Это можно сравнить со спектром гамильтониана полной энергии квантовой системы. Распространение волн идентификаторов можно сопоставить с эволюцией волновой функции. Описание, которое несет идентификатор, переходит в активность паттернов. Далее мы покажем, что при этом активируются не все паттерны, а только те, что могут описать непротиворечивую картину, соответствующую контексту происходящего. В этот момент наша система переходит в одно из возможных разрешенных состояний, что сильно напоминает коллапс волновой функции.
Использованная литература
Продолжение
Предыдущие части:
Часть 1. Нейрон
Часть 2. Факторы
Часть 3. Персептрон, сверточные сети
Часть 4. Фоновая активность
Часть 5. Волны мозга
Часть 6. Система проекций
Часть 7. Интерфейс человек-компьютер
Часть 8. Выделение факторов в волновых сетях
Часть 9. Паттерны нейронов-детекторов. Обратная проекция
Алексей Редозубов (2014)
